The Flagellum Unspun
The Collapse of "Irreducible Complexity"
Kenneth R. Miller
Brown University
Providence, Rhode Island 02912 USA
This
is a pre-publication copy of an article that
will appear in volume entitled "Debating Design:
from Darwin to DNA," edited by Michael Ruse and
William Dembski, which will be published by
Cambridge University Press volume in 2004.
Almost from the moment The Origin
of Species was published in 1859, the opponents of evolution
have fought a long, losing battle against their Darwinian foes.
Today, like a prizefighter in the late rounds losing badly on
points, they've placed their hopes in one big punch – a single
claim that might smash through the overwhelming weight of
scientific evidence to bring Darwin to the canvas once and for
all. Their name for this virtual roundhouse right is
"intelligent design."
In the last several years, the
intelligent design movement has attempted to move against
science education standards in several American states, most
famously in Kansas and Ohio (Holden 1999; Gura 2002). The
principal claim made by adherents of this view is that they can
detect the presence of "intelligent design" in complex
biological systems. As evidence, they cite a number of specific
examples, including the vertebrate blood clotting cascade, the
eukaryotic cilium, and most notably, the eubacterial flagellum (Behe
1996a, Behe 2002).
Of all these examples, the flagellum
has been presented so often as a counter-example to evolution
that it might well be considered the "poster child" of the
modern anti-evolution movement. Variations of its image (Figure
1) now appear on web pages of anti-evolution groups like the
Discovery Institute, and on the covers of "intelligent design"
books such as William Dembski's No Free Lunch (Dembski
2002a). To anti-evolutionists, the high status of the flagellum
reflects the supposed fact that it could not possibly have been
produced by an evolutionary pathway.
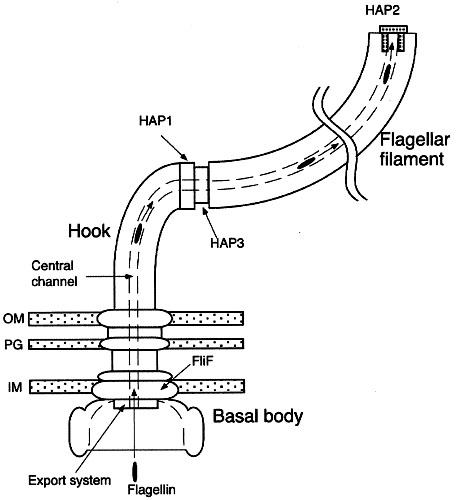
Figure 1: The
eubacterial flagellum. The flagellum is an ion-powered
rotary motor, anchored in the membranes surrounding the
bacterial cell. This schematic diagram highlights the
assembly process of the bacterial flagellar filament and
the cap-filament complex. OM, outer membrane; PG,
peptidoglycan layer; IM, cytoplasmic membrane (From
Yonekura et al 2000).
There is, to be sure, nothing new or
novel in an anti-evolutionist pointing to a complex or intricate
natural structure, and professing skepticism that it could have
been produced by the "random" processes of mutation and natural
selection. Nonetheless, the "argument from personal
incredulity," as such sentiment has been appropriately
described, has been a weapon of little value in the
anti-evolution movement. Anyone can state at any time that
they cannot imagine how evolutionary mechanisms might have
produced a certain species, organ, structure. Such statements,
obviously, are personal – and they say more about the
limitations of those who make them than they do about the
limitations of Darwinian mechanisms.
The hallmark of the intelligent design
movement, however, is that it purports to rise above the level
of personal skepticism. It claims to have found a reason
why evolution could not have produced a structure like the
bacterial flagellum, a reason based on sound, solid scientific
evidence.
Why does the intelligent design
movement regard the flagellum as unevolvable? Because it is said
to possesses a quality known as "irreducible complexity."
Irreducibly complex structures, we are told, could not have been
produced by evolution, or, for that matter, by any natural
process. They do exist, however, and therefore they must have
been produced by something. That something could only be an
outside intelligent agency operating beyond the laws of nature –
an intelligent designer. That, simply stated, is the core of the
new argument from design, and the intellectual basis of the
intelligent design movement.
The great irony of the flagellum's
increasing acceptance as an icon of anti-evolution is that fact
that research had demolished its status as an example of
irreducible complexity almost at the very moment it was first
proclaimed. The purpose of this article is to explore the
arguments by which the flagellum's notoriety has been achieved,
and to review the research developments that have now undermined
they very foundations of those arguments.
The Argument's Origins
The flagellum owes its status
principally to Darwin's Black Box (Behe 1996a) a book by
Michael Behe that employed it in a carefully-crafted
anti-evolution argument. Building upon William Paley's
well-known "argument from design," Behe sought to bring the
argument two centuries forward into the realm of biochemistry.
Like Paley, Behe appealed to his readers to appreciate the
intricate complexity of living organisms as evidence for the
work of a designer. Unlike Paley, however, he raised the
argument to a new level, claiming to have discovered a
scientific principle that could be used to prove that certain
structures could not have been produced by evolution. That
principle goes by the name of "irreducible complexity."
An irreducibly complex structure is
defined as ". . . a single system composed of several
well-matched, interacting parts that contribute to the basic
function, wherein the removal of any one of the parts causes the
system to effectively cease functioning." (Behe 1996a, 39) Why
would such systems present difficulties for Darwinism? Because
they could not possibly have been produced by the process of
evolution:
"An irreducibly complex
system cannot be produced directly by numerous,
successive, slight modifications of a precursor system,
because any precursor to an irreducibly complex system
that is missing a part is by definition nonfunctional.
.... Since natural selection can only choose systems
that are already working, then if a biological system
cannot be produced gradually it would have to arise as
an integrated unit, in one fell swoop, for natural
selection to have anything to act on." (Behe 1996b)
The phrase "numerous, successive,
slight modifications" is not accidental. The very same words
were used by Charles Darwin in The Origin of Species in
describing the conditions that had to be met for his theory to
be true. As Darwin wrote, if one could find an organ or
structure that could not have been formed by "numerous,
successive, slight modifications," his "theory would absolutely
break down" (Darwin 1859, 191). To anti-evolutionists, the
bacterial flagellum is now regarded as exactly such a case – an
"irreducibly complex system" which "cannot be produced directly
by numerous successive, slight modifications." A system that
could not have evolved – a desperation punch that just might win
the fight in the final round – a tool with which the theory of
evolution can be brought down.
The Logic of Irreducible
Complexity
Living cells are filled, of course,
with complex structures whose detailed evolutionary origins are
not known. Therefore, in fashioning an argument against
evolution one might pick nearly any cellular structure, the
ribosome for example, and claim – correctly – that its origin
has not been explained in detail by evolution.
Such arguments are easy to make, of
course, but nature of scientific progress renders them far from
compelling. The lack of a detailed current explanation for a
structure, organ, or process does not mean that science will
never come up with one. As an example, one might consider the
question of how left-right asymmetry arises in vertebrate
development, a question that was beyond explanation until the
1990s (Belmonte 1999). In 1990 one might have argued that the
body's left-right asymmetry could just as well be explained by
the intervention of a designer as by an unknown molecular
mechanism. Only a decade later, the actual molecular mechanism
was identified (Stern 2002), and any claim one might have made
for the intervention of a designer would have been discarded.
The same point can be made, of course, regarding any structure
or mechanism whose origins are not yet understood.
The utility of the bacterial flagellum
is that it seems to rise above this "argument from ignorance."
By asserting that it is a structure "in which the removal of an
element would cause the whole system to cease functioning" (Behe
2002), the flagellum is presented as a "molecular machine" whose
individual parts must have been specifically crafted to work as
a unified assembly. The existence of such a multipart machine
therefore provides genuine scientific proof of the actions of an
intelligent designer.
In the case of the flagellum, the
assertion of irreducible complexity means that a minimum number
of protein components, perhaps 30, are required to produce a
working biological function. By the logic of irreducible
complexity, these individual components should have no function
until all 30 are put into place, at which point the function of
motility appears. What this means, of course, is that evolution
could not have fashioned those components a few at a time, since
they do not have functions that could be favored by natural
selection. As Behe wrote: " . . . natural selection can only
choose among systems that are already working" (Behe 2002), and
an irreducibly complex system does not work unless all of its
parts are in place. The flagellum is irreducibly complex, and
therefore, it must have been designed. Case closed.
Answering the Argument
The assertion that cellular machines
are irreducibly complex, and therefore provide proof of design,
has not gone unnoticed by the scientific community. A number of
detailed rebuttals have appeared in the literature, and many
have pointed out the poor reasoning of recasting the classic
argument from design in the modern language of biochemistry
(Coyne 1996; Miller 1996; Depew 1998; Thornhill and Ussery
2000). I have suggested elsewhere that the scientific literature
contains counter-examples to any assertion that evolution cannot
explain biochemical complexity (Miller 1999, 147), and other
workers have addressed the issue of how evolutionary mechanisms
allow biological systems to increase in information content
(Schneider 2000; Adami, Ofria, and Collier 2000).
The most powerful rebuttals to the
flagellum story, however, have not come from direct attempts to
answer the critics of evolution. Rather, they have emerged from
the steady progress of scientific work on the genes and proteins
associated with the flagellum and other cellular structures.
Such studies have now established that the entire premise by
which this molecular machine has been advanced as an argument
against evolution is wrong – the bacterial flagellum is not
irreducibly complex. As we will see, the flagellum – the
supreme example of the power of this new "science of design" –
has failed its most basic scientific test. Remember the claim
that "any precursor to an irreducibly complex system that is
missing a part is by definition nonfunctional?" As the evidence
has shown, nature is filled with examples of "precursors" to the
flagellum that are indeed "missing a part," and yet are
fully-functional. Functional enough, in some cases, to pose a
serious threat to human life.
The Type -III Secretory
Apparatus
In the popular imagination, bacteria
are "germs" – tiny microscopic bugs that make us sick.
Microbiologists smile at that generalization, knowing that most
bacteria are perfectly benign, and many are beneficial – even
essential – to human life. Nonetheless, there are indeed
bacteria that produce diseases, ranging from the mildly
unpleasant to the truly dangerous. Pathogenic, or
disease-causing, bacteria threaten the organisms they infect in
a variety of ways, one of which is to produce poisons and inject
them directly into the cells of the body. Once inside, these
toxins break down and destroy the host cells, producing illness,
tissue damage, and sometimes even death.
In order to carry out this diabolical
work, bacteria must not only produce the protein toxins that
bring about the demise of their hosts, but they must efficiently
inject them across the cell membranes and into the cells of
their hosts. They do this by means of any number of specialized
protein secretory systems. One, known as the type III secretory
system (TTSS), allows gram negative bacteria to translocate
proteins directly into the cytoplasm of a host cell (Heuck
1998). The proteins transferred through the TTSS include a
variety of truly dangerous molecules, some of which are known as
"virulence factors," and are directly responsible for the
pathogenic activity of some of the most deadly bacteria in
existence (Büttner and Bonas 2002; Heuck 1998).
At first glance, the existence of the
TTSS, a nasty little device that allows bacteria to inject these
toxins through the cell membranes of its unsuspecting hosts,
would seem to have little to do with the flagellum. However,
molecular studies of proteins in the TTSS have revealed a
surprising fact – the proteins of the TTSS are directly
homologous to the proteins in the basal portion of the bacterial
flagellum. As figure 2 (Heuck 1998) shows, these homologies
extend to a cluster of closely-associated proteins found in both
of these molecular "machines." On the basis of these homologies,
McNab (McNab 1999) has argued that the flagellum itself should
be regarded as a type III secretory system. Extending such
studies with a detailed comparison of the proteins associated
with both systems, Aizawa has seconded this suggestion, noting
that the two systems "consist of homologous component proteins
with common physico-chemical properties" (Aizawa 2001, 163). It
is now clear, therefore, that a smaller subset of the full
complement of proteins in the flagellum makes up the functional
transmembrane portion of the TTSS.
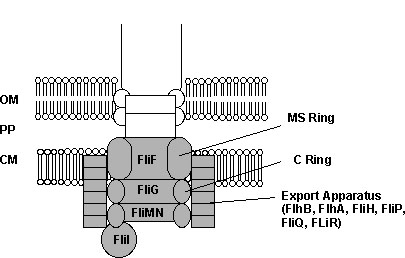
Figure 2: There
are extensive homologies between type III secretory
proteins and proteins involved in export in the basal
region of the bacterial flagellum. These homologies
demonstrate that the bacterial flagellum is not
"irreducibly complex." In this diagram (redrawn from
Heuck 1998), the shaded portions of the basal region
indicate proteins in the E. coli flagellum
homologous to the Type III secretory structure of
Yersinia. . OM, outer membrane; PP, periplasmic
space; CM, cytoplasmic membrane.
Stated directly, the TTSS does its
dirty work using a handful of proteins from the base of the
flagellum. From the evolutionary point of view, this
relationship is hardly surprising. In fact, it's to be expected
that the opportunism of evolutionary processes would mix and
match proteins to produce new and novel functions. According to
the doctrine of irreducible complexity, however, this should not
be possible. If the flagellum is indeed irreducibly complex,
then removing just one part, let alone 10 or 15, should render
what remains "by definition nonfunctional." Yet the TTSS is
indeed fully-functional, even though it is missing most of the
parts of the flagellum. The TTSS may be bad news for us, but for
the bacteria that possess it, it is a truly valuable biochemical
machine.
The existence of the TTSS in a wide
variety of bacteria demonstrates that a small portion of the
"irreducibly complex" flagellum can indeed carry out an
important biological function. Since such a function is clearly
favored by natural selection, the contention that the flagellum
must be fully-assembled before any of its component parts can be
useful is obviously incorrect. What this means is that the
argument for intelligent design of the flagellum has failed.
Counterattack
Classically, one of the most
widely-repeated charges made by anti-evolutionists is that the
fossil record contains wide "gaps" for which transitional
fossils have never been found. Therefore, the intervention of a
creative agency, an intelligent designer, must be invoked to
account for each gap. Such gaps, of course, have been filled
with increasing frequency by paleontologists – the increasingly
rich fossil sequences demonstrating the origins of whales are a
useful examples (Thewissen, Hussain, and Arif 1994; Thewissen,
Williams, Roe, and Hussain 2001). Ironically, the response of
anti-evolutionists to such discoveries is frequently to claim
that things have only gotten worse for evolution. Where
previously there had been just one gap, as a result of the
transitional fossil, now there are two (one on either side of
the newly-discovered specimen).
As word of the relationship between the
eubacterial flagellum and the TTSS has begun to spread among the
"design" community, the first hints of a remarkably similar
reaction have emerged. The TTSS only makes problems worse for
evolution, according to this response, because now there are two
irreducibly-complex systems to deal with. The flagellum is still
irreducibly complex – but so is the TTSS. But now there are two
systems for evolutionists to explain instead of just one.
Unfortunately for this line of
argument, the claim that one irreducibly-complex system might
contain another is self-contradictory. To understand this, we
need to remember that the entire point of the design argument,
as exemplified by the flagellum, is that only the entire
biochemical machine, with all of its parts, is functional. For
the intelligent design argument to stand, this must be the case,
since it provides the basis for their claim that only the
complete flagellum can be favored by natural selection, not any
its component parts.
However, if the flagellum contains
within it a smaller functional set of components like the TTSS,
then the flagellum itself cannot be irreducibly complex – by
definition. Since we now know that this is indeed the case, it
is obviously true that the flagellum is not irreducibly complex.
A second reaction, which I have heard
directly after describing the relationship between the secretory
apparatus and the flagellum, is the objection that the TTSS does
not tell us how either it or the flagellum evolved. This is
certainly true, although Aizawa has suggested that the TTSS may
indeed be an evolutionary precursor of the flagellum (Aizawa
2001). Nonetheless, until we have produced a step-by-step
account for the evolutionary derivation of the flagellum, one
may indeed invoke the argument from ignorance for this and every
other complex biochemical machine.
However, in agreeing to this, one must
keep in mind that the doctrine of irreducible complexity was
intended to go one step beyond the claim of ignorance. It was
fashioned in order to provide a rationale for claiming that the
bacterial flagellum couldn't have evolved, even in principle,
because it is irreducibly complex. Now that a simpler,
functional system (the TTSS) has been discovered among the
protein components of the flagellum, the claim of irreducible
complexity has collapsed, and with it any "evidence" that the
flagellum was designed.
Combinatorial Argument
At first glance, William Dembski's case
for intelligent design seems to follow a distinctly different
strategy in dealing with biological complexity. His recent book,
No Free Lunch (Dembski 2002a), lays out this case, using
information theory and mathematics to show that life is the
result of intelligent design. Dembski makes the assertion that
living organisms contain what he calls "complex specified
information" (CSI), and claims to have shown that the
evolutionary mechanism of natural selection cannot produce CSI.
Therefore, any instance of CSI in a living organism must be the
result of intelligent design. And living organisms, according to
Dembski, are chock-full of CSI.
Dembski's arguments, couched in the
language of information theory, are highly technical and are
defended, almost exclusively, by reference to their utility in
detecting information produced by human beings. These include
phone and credit card numbers, symphonies, and artistic
woodcuts, to name just a few. One might then expect that
Dembski, having shown how the presence of CSI can be
demonstrated in man made objects, would then turn to a variety
of biological objects. Instead, he turns to just one such
object, the bacterial flagellum.
Dembski then offers his readers a
calculation showing that the flagellum could not have possibly
have evolved. Significantly, he begins that calculation by
linking his arguments to those of Behe, writing: "I want
therefore in this section to show how irreducible complexity is
a special case of specified complexity, and in particular I want
to sketch how one calculates the relevant probabilities needed
to eliminate chance and infer design for such systems" (Dembski
2002a, 289). Dembski then tells us that an irreducibly complex
system, like the flagellum, is a "discrete combinatorial
object." What this means, as he explains, is that the
probability of assembling such an object can be calculated by
determining the probabilities that each of its components might
have originated by chance, that they might have been localized
to the same region of the cell, and that they would be assembled
in precisely the right order. Dembski refers to these three
probabilities as Porig,
Plocal,
and Pconfig,
and he regards each of them as separate and independent (Dembski
2002a, 291).
This approach overlooks the fact that
the last two probabilities are actually contained within the
first. Localization and self-assembly of complex protein
structures in prokaryotic cells are properties generally
determined by signals built into the primary structures of the
proteins themselves. The same is likely true for the amino acid
sequences of the 30 or so protein components of the flagellum
and the approximately 20 proteins involved in the flagellum's
assembly (McNab 1999; Yonekura et al 2000). Therefore, if
one gets the sequences of all the proteins right, localization
and assembly will take care of themselves.
To the ID enthusiast, however, this is
a point of little concern. According to Dembski, evolution could
still not construct the 30 proteins needed for the flagellum.
His reason is that the probability of their assembly falls below
what he terms the "universal probability bound." According to
Dembski, the probability bound is a sensible allowance for the
fact that highly improbable events do occur from time to time in
nature. To allow for such events, he agrees that given enough
time, any event with a probability larger than 10-150
might well take place. Therefore, if a sequence of events, such
as a presumed evolutionary pathway, has a calculated probability
less than 10-150
, we may conclude that the pathway is impossible. If the
calculated probability is greater than 10-150,
it's possible (even if unlikely).
When Dembski turns his attention to the
chances of evolving the 30 proteins of the bacterial flagellum,
he makes what he regards as a generous assumption. Guessing that
each of the proteins of the flagellum have about 300 amino
acids, one might calculate that the chances of getting just one
such protein to assemble from "random" evolutionary processes
would be 20-300
, since there are 20 amino acids specified by the genetic code.
Dembski, however, concedes that proteins need not get the
exact amino acid sequence right in order to be functional,
so he cuts the odds to just 20-30,
which he tells his readers is "on the order of 10-39"
(Dembski 2002a, 301). Since the
flagellum requires 30 such proteins, he explains that 30 such
probabilities "will all need to be multiplied to form the
origination probability"(Dembski 2002a, 301). That would give us
an origination probability for the flagellum of 10
-1170,
far below the universal probability bound. The flagellum
couldn't have evolved, and now we have the numbers to prove it.
Right?
Assuming Impossibility
I have no doubt that to the casual
reader, a quick glance over the pages of numbers and symbols in
Dembski's books is impressive, if not downright intimidating.
Nonetheless, the way in which he calculates the probability of
an evolutionary origin for the flagellum shows how little
biology actually stands behind those numbers. His computation
calculates only the probability of spontaneous, random assembly
for each of the proteins of the flagellum. Having come up with a
probability value on the order of 10
-1170,
he assures us that he has shown the flagellum to be unevolvable.
This conclusion, of course, fits comfortably with his view is
that "The Darwinian mechanism is powerless to produce
irreducibly complex systems..." (Dembski 2002a, 289).
However complex Dembski's analysis, the
scientific problem with his calculations is almost too easy to
spot. By treating the flagellum as a "discrete combinatorial
object" he has shown only that it is unlikely that the parts
flagellum could assemble spontaneously. Unfortunately for his
argument, no scientist has ever proposed that the flagellum or
any other complex object evolved that way. Dembski, therefore,
has constructed a classic "straw man" and blown it away with an
irrelevant calculation.
By treating the flagellum as a discrete
combinatorial object he has assumed in his calculation that no
subset of the 30 or so proteins of the flagellum could have
biological activity. As we have already seen, this is wrong.
Nearly a third of those proteins are closely related to
components of the TTSS, which does indeed have biological
activity. A calculation that ignores that fact has no scientific
validity.
More importantly, Dembski's willingness
to ignore the TTSS lays bare the underlying assumption of his
entire approach towards the calculation of probabilities and the
detection of "design." He assumes what he is trying to prove.
According to Dembski, the detection of
"design" requires that an object display complexity that could
not be produced by what he calls "natural causes." In order to
do that, one must first examine all of the possibilities by
which an object, like the flagellum, might have been generated
naturally. Dembski and Behe, of course, come to the conclusion
that there are no such natural causes. But how did they
determine that? What is the scientific method used to support
such a conclusion? Could it be that their assertions of the lack
of natural causes simply amount to an unsupported personal
belief? Suppose that there are such causes, but they simply
happened not to think of them? Dembski actually seems to realize
that this is a serious problem. He writes: "Now it can happen
that we may not know enough to determine all the relevant chance
hypotheses [which here, as noted above, means all relevant
natural processes (hvt)]. Alternatively, we might think we
know the relevant chance hypotheses, but later discover that we
missed a crucial one. In the one case a design inference could
not even get going; in the other, it would be mistaken" (Dembski
2002, 123 (note 80)).
What Dembski is telling us is that in
order to "detect" design in a biological object one must first
come to the conclusion that the object could not have been
produced by any "relevant chance hypotheses" (meaning,
naturally, evolution). Then, and only then, are Dembski's
calculations brought into play. Stated more bluntly, what this
really means is that the "method" first involves assuming the
absence of an evolutionary pathway leading to the object,
followed by a calculation "proving" the impossibility of
spontaneous assembly. Incredibly, this a priori reasoning
is exactly the sort of logic upon which the new "science of
design" has been constructed.
Not surprisingly, scientific reviewers
have not missed this point – Dembski's arguments have been
repeatedly criticized on this issue and on many others (Orr
2002; Charlesworth 2002; Padian 2002).
Designing the Cycle
In assessing the design argument,
therefore, it only seems as though two distinct arguments
have been raised for the unevolvability of the flagellum. In
reality, those two arguments, one invoking irreducible
complexity and the other specified complex information, both
depend upon a single scientifically insupportable position.
Namely, that we can look at a complex biological object and
determine with absolute certainty that none of its component
parts could have been first selected to perform other functions.
The discovery of extensive homologies between the Type III
secretory system and the flagellum has now shown just how wrong
that position was.
When anti-evolutionary arguments
featuring the bacterial flagellum rose into prominence,
beginning with the 1996 publication of Darwin's Black Box (Behe
1996a), they were predicated upon the assertion that each of the
protein components of the flagellum were crafted, in a single
act of design, to fit the specific purpose of the flagellum. The
flagellum was said to be unevolvable since the entire complex
system had to be assembled first in order to produce any
selectable biological function. This claim was broadened to
include all complex biological systems, and asserted further
that science would never find an evolutionary pathway to any of
these systems. After all, it hadn't so far, at least according
to one of "design's" principal advocates:
There is no publication in
the scientific literature – in prestigious journals,
specialty journals, or books – that describes how
molecular evolution of any real, complex, biochemical
system either did occur or even might have occurred.
(Behe 1996a, 185)
As many critics of intelligent design
have pointed out, that statement is simply false. Consider, as
just one example, the Krebs cycle, an intricate biochemical
pathway consisting of nine enzymes and a number of cofactors
that occupies center stage in the pathways of cellular
metabolism. The Krebs cycle is "real," "complex," and
"biochemical." Does it also present a problem for evolution?
Apparently yes, according to the authors of a 1996 paper in the
Journal of Molecular evolution, who wrote:
"The Krebs cycle has been
frequently quoted as a key problem in the evolution of
living cells, hard to explain by Darwin’s natural
selection: How could natural selection explain the
building of a complicated structure in toto, when the
intermediate stages have no obvious fitness
functionality? (Melendez-Hevia, Wadell, and Cascante
1996)
Where intelligent design theorists
throw up their hands and declare defeat for evolution, however,
these researchers decided to do the hard scientific work of
analyzing the components of the cycle, and seeing if any of them
might have been selected for other biochemical tasks. What they
found should be a lesson to anyone who asserts that evolution
can only act by direct selection for a final function. In fact,
nearly all of the proteins of the complex cycle can serve
different biochemical purposes within the cell, making it
possible to explain in detail how they evolved:
In the Krebs cycle problem
the intermediary stages were also useful, but for
different purposes, and, therefore, its complete design
was a very clear case of opportunism. . . . the Krebs
cycle was built through the process that Jacob (1977)
called ‘‘evolution by molecular tinkering,’’ stating
that evolution does not produce novelties from scratch:
It works on what already exists. The most novel result
of our analysis is seeing how, with minimal new
material, evolution created the most important pathway
of metabolism, achieving the best chemically possible
design. In this case, a chemical engineer who was
looking for the best design of the process could not
have found a better design than the cycle which works in
living cells." (Melendez-Hevia, Wadell, and Cascante
1996)
Since this paper appeared, a study
based on genomic DNA sequences has confirmed the validity of
this approach (Huynen, Dandekar, and Bork 1999). By contrast,
how would intelligent design have approached the Krebs Cycle?
Using Dembski's calculations as our guide, we would first
determine the amino acid sequences of each of the proteins of
the cycle, and then calculate the probability of their
spontaneous assembly. When this is done, an origination
probability of less than 10
-400
is the result. Therefore, the result of applying "design" as a
predictive science would have told both groups of researchers
that their ultimately successful studies would have been
fruitless, since the probability of spontaneous assembly falls
below the "universal probability bound."
We already know, however, the reason
that such calculations fail. They carry a built-in assumption
that the component parts of a complex biochemical system have no
possible functions beyond the completely assembled system
itself. As we have seen, this assumption is false. The Krebs
cycle researchers knew better, of course, and were able to
produce two important studies describing how a real, complex,
biochemical system might have evolved – the very thing that
design theorists once claimed did not exist in the scientific
literature.
The Failure of Design
It is no secret that concepts like
"irreducible complexity" and "intelligent design" have failed to
take the scientific community by storm (Forrest 2002). Design
has not prompted new research studies, new breakthroughs, or
novel insights on so much as a single scientific question.
Design advocates acknowledge this from time to time, but they
often claim that this is because the scientific deck is stacked
against them. The Darwinist establishment, they say, prevents
them from getting a foot in the laboratory door.
I would suggest that the real reason
for the cold shoulder given "design" by the scientific
community, particularly by life science researchers, is because
time and time again its principal scientific claims have turned
out to be wrong. Science is a pragmatic activity, and if your
hypothesis doesn't work, it is quickly discarded.
The claim of irreducible complexity for
the bacterial flagellum is an obvious example of this, but there
are many others. Consider, for example, the intricate cascade of
proteins involved in the clotting of vertebrate blood. This has
been cited as one of the principal examples of the kind of
complexity that evolution cannot generate, despite the elegant
work of Russell Doolittle (Doolittle and Feng 1987; Doolittle
1993) to the contrary. A number of proteins are involved in this
complex pathway, as described by Behe:
When an animal is cut, a
protein called Hagemann factor (XII) sticks to the
surface of cells near the wound. Bound Hagemann factor
is then cleaved by a protein called HMK to yield
activated Hagemann factor. Immediately the activated
Hagemann factor converts another protein, called
prekallikrein, to its active form, kallikrein. (Behe
1996a, 84)
How important are each of these
proteins? In line with the dogma of irreducible complexity, Behe
argues that each and every component must be in place before the
system will work, and he is perfectly clear on this point:
. . . none of the cascade
proteins are used for anything except controlling the
formation of a clot. Yet in the absence of any of the
components, blood does not clot, and the system fails.
(Behe 1996a, 86)
As we have seen, the claim that every
one of the components must be present for clotting to work is
central to the "evidence" for design. One of those components,
as these quotations indicate, is Factor XII, which initiates the
cascade. Once again, however, a nasty little fact gets in the
way of intelligent design theory. Dolphins lack Factor XII
(Robinson, Kasting, and Aggeler 1969), and yet their blood clots
perfectly well. How can this be if the clotting cascade is
indeed irreducibly complex? It cannot, of course, and therefore
the claim of irreducible complexity is wrong for this system as
well. I would suggest, therefore, that the real reason for the
rejection of "design" by the scientific community is remarkably
simple – the claims of the intelligent design movement are
contradicted time and time again by the scientific evidence.
The Flagellum Unspun
In any discussion of the question of
"intelligent design," it is absolutely essential to determine
what is meant by the term itself. If, for example, the advocates
of design wish to suggest that the intricacies of nature, life,
and the universe reveal a world of meaning and purpose
consistent with an overarching, possibly Divine intelligence,
then their point is philosophical, not scientific. It is a
philosophical point of view, incidentally, that I share, along
with many scientists. As H. Allen Orr pointed out in a recent
review:
Plenty of scientists have,
after all, been attracted to the notion that natural
laws reflect (in some way that's necessarily poorly
articulated) an intelligence or aesthetic sensibility.
This is the religion of Einstein, who spoke of "the
grandeur of reason incarnate in existence" and of the
scientist's "religious feeling [that] takes the form of
a rapturous amazement at the harmony of natural law."
(Orr 2002).
This, however, is not what is meant by
"intelligent design" in the parlance of the new
anti-evolutionists. Their views demand not a universe in which
the beauty and harmony of natural law has brought a world of
vibrant and fruitful life into existence, but rather a universe
in which the emergence and evolution of life is made expressly
impossible by the very same rules. Their view requires that the
source of each and every novelty of life was the direct and
active involvement of an outside designer whose work violated
the very laws of nature he had fashioned. The world of
intelligent design is not the bright and innovative world of
life that we have come to know through science. Rather, it is a
brittle and unchanging landscape, frozen in form and unable to
adapt except at the whims of its designer.
Certainly, the issue of design and
purpose in nature is a philosophical one that scientists can and
should discuss with great vigor. However, the notion at the
heart's of today intelligent design movement is that the direct
intervention of an outside designer can be demonstrated by the
very existence of complex biochemical systems. What even they
acknowledge is that their entire scientific position rests upon
a single assertion – that the living cell contains biochemical
machines that are irreducibly complex. And the bacterial
flagellum is the prime example of such a machine.
Such an assertion, as we have seen, can
be put to the test in a very direct way. If we are able to
search and find an example of a machine with fewer protein
parts, contained within the flagellum, that serves a purpose
distinct from motility, the claim of irreducible complexity is
refuted. As we have also seen, the flagellum does indeed contain
such a machine, a protein-secreting apparatus that carries out
an important function even in species that lack the flagellum
altogether. A scientific idea rises or falls on the weight of
the evidence, and the evidence in the case of the bacterial
flagellum is abundantly clear.
As an icon of anti-evolution, the
flagellum has fallen.
The very existence of the Type III
Secretory System shows that the bacterial flagellum is not
irreducibly complex. It also demonstrates, more generally, that
the claim of "irreducible complexity" is scientifically
meaningless, constructed as it is upon the flimsiest of
foundations – the assertion that because science has not yet
found selectable functions for the components of a certain
structure, it never will. In the final analysis, as the claims
of intelligent design fall by the wayside, its advocates are
left with a single, remaining tool with which to battle against
the rising tide of scientific evidence. That tool may be
effective in some circles, of course, but the scientific
community will be quick to recognize it for what it really is –
the classic argument from ignorance, dressed up in the shiny
cloth of biochemistry and information theory.
When three leading advocates of
intelligent design were recently given a chance to make their
case in an issue of Natural History magazine, they each
concluded their articles with a plea for design. One wrote that
we should recognize "the design inherent in life and the
universe" (Behe 2002), another that "design remains a
possibility" (Wells 2002), and another "that the natural
sciences need to leave room for design" (Dembski 2002b). Yes, it
is true. Design does remain a possibility, but not the type of
"intelligent design" of which they speak.
As Darwin wrote, there is grandeur in
an evolutionary view of life, a grandeur that is there for all
to see, regardless of their philosophical views on the meaning
and purpose of life. I do not believe, even for an instant, that
Darwin's vision has weakened or diminished the sense of wonder
and awe that one should feel in confronting the magnificence and
diversity of the living world. Rather, to a person of faith it
should enhance their sense of the Creator's majesty and wisdom
(Miller 1999). Against such a backdrop, the struggles of the
intelligent design movement are best understood as clamorous and
disappointing double failures – rejected by science because they
do not fit the facts, and having failed religion because they
think too little of God.
Bibliography
Adami, C., C. Ofria, and T. C.
Collier, 2000. Evolution of biological complexity,
Proceedings of the National Academy of Sciences 97:
4463—4468.
Aizawa, S.-I., 2001.
Bacterial flagella and type III secretion systems,
FEMS Microbiology Letters 202: 157-164.
Behe, M, 1996b. Evidence for
Intelligent Design from Biochemistry, a speech given at
the Discovery Institute's God & Culture Conference,
August 10, 1996 Seattle, WA. Available from World Wide
Web: http://www.arn.org/docs/behe/mb_idfrombiochemistry.htm
Behe, M. 1996a. Darwin's
Black Box. New York: The Free Press.
Behe, M., 2002. The challenge
of irreducible complexity, Natural History 111
(April): 74.
Büttner D., and U. Bonas, 2002.
Port of entry - the Type III secretion translocon,
Trends in Microbiology 10: 186-191.
Belmonte, J. C. I. , 1999. How
the body tells right from left. Scientific American
280 (June): 46-51.
Charlesworth, B., 2002.
Evolution by Design? Nature 418: 129.
Coyne, J. A., 1996. God in the
details, Nature 383: 227-228.
Darwin, C. 1872. The Origin
of Species (6th edition). London: Oxford University
Press.
Dembski, W. 2002a, No Free
Lunch: Why specified complexity cannot be purchased
without intelligence. Lanham, Maryland: Rowman &
Littlefield.
Dembski, W., 2002b. Detecting
design in the natural sciences, Natural History
111 (April): 76.
Depew, D. J. (1998),
Intelligent design and irreducible complexity: A
rejoinder. Rhetoric and Public Affairs 1:
571-578.
Doolittle, R. F., 1993. The
evolution of vertebrate blood coagulation: A case of yin
and yang, Thrombosis and Heamostasis 70: 24-28.
Doolittle, R. F., and D. F.
Feng, 1987. Reconstructing the evolution of vertebrate
blood coagulation from a consideration of the amino acid
sequences of clotting proteins. Cold Spring Harbor
Symposia on Quantitative Biology 52: 869-874.
Forrest B., 2002. The newest
evolution of creationism. Natural History 111
(April): 80.
Gura, T., 2002, Evolution
Critics seek role for unseen hand in education.
Nature 416: 250.
Heuck, C. J., 1998. Type III
protein secretion systems in bacterial pathogens of
animals and plants, Microbiol. Mol. Biol. Rev.
62: 379-433.
Holden, C., 1999. Kansas Dumps
Darwin, Raises Alarm Across the United States.
Science 285: 1186-1187.
Huynen, M. A., T. Dandekar, and
P. Bork (1999) Variation and evolution of the
citric-acid cycle: a genomic perspective. Trends in
Microbiology 7: 281-291.
McNab, R. M., 1999. The
Bacterial Flagellum: Reversible Rotary Propellor and
Type III Export Apparatus. Journal of Bacteriology
181: 7149—7153.
Melendez-Hevia, E., T. G.
Wadell, and M. Cascante, 1996. The puzzle of the Krebs
citric acid cycle: Assembling the pieces of chemically
feasible reactions, and opportunism in the design of
metabolic pathways during evolution. J. Molecular
Evolution 43: 293—303
Miller, K. R. 1999. Finding
Darwin's God. New York: Harper Collins.
Miller, K. R., 1996. A review
of Darwin's Black Box, Creation/Evolution 16:
36-40.
Orr, H. A., 2002. The Return of
Intelligent Design, The Boston Review (Summer
2002 Issue).
Padian, K., 2002. Waiting for
the Watchmaker, Science 295: 2373-2374.
Pennock, R. T., 2001.
Intelligent Design Creationism and Its Critics:
Philosophical, Theological, and Scientific Perspectives.
Cambridge, Massachusetts: MIT Press.
Robinson, A. J., M. Kropatkin,
and P. M. Aggeler, 1969. Hagemann Factor (Factor XII)
Deficiency in Marine Mammals. Science 166:
1420-1422.
Schneider, T.D. (2000),
Evolution of biological information. Nucleic Acids
Research 28: 2794-2799.
Stern, C., 2002. Embryology:
Fluid flow and broken symmetry, Nature 418:
29-30.
Thewissen, J. G, M., E. M.
Williams, L. J. Roe, and S. T. Hussain, 2001. Skeletons
of terrestrial cetaceans and the relationship of whales
to artiodactyls, Nature 413: 277-281.
Thewissen, J. G. M., S. T.
Hussain, and M. Arif, 1994. Fossil evidence for the
origin of aquatic locomotion in archaeocete whales,
Science 286: 210-212.
Thornhill, R. H., and D. W. and
Ussery, 2000. A classification of possible routes of
Darwinian evolution, The Journal of Theoretical
Biology 203: 111-116.
Wells, J, 2002. Elusive Icons
of Evolution, Natural History 111 (April): 78.
Yonekura, K., S. Maki, D. G.
Morgan, D. J. DeRosier, F.Vonderviszt, K.Imada, and K.
Namba, 2000. The Bacterial Flagellar Cap as the Rotary
Promoter of Flagellin Self-Assembly, Science 290:
2148-2152.
|